
Table des matières
Note sur la dormance des graines
Type de documentation
Cette page est une documentation en forme d’explication.
Vous pouvez partager vos connaissances en l’améliorant (comment ?).
Cette page est axée sur la compréhension, explique, fournit des renseignements généraux et le contexte. Elle est comparable à un article sur l’histoire sociale de la tomate ou l’histoire sociale culinaire.
Exemple : Le wiki de communs
Répertoire : Les explications dans ce wiki
Support : Le portail dédié à la documentation et aux codes sources
Dans le champs de la fr:dormance des graines il y a 4 grandes figures de proues du travail scientifique :
- Lela Viola Barton
- Marianna Georgievna Nikolaeva
- Carol C. Baskin
- Jerry M. Baskin
Nous prendrons ici un appui de départ depuis Seeds. Ecology, Biogeography, and, Evolution of Dormancy and Germination Academic Press, 2, 2014 Carol C. Baskin and Jerry M. Baskin.
Dans cet ouvrage, Baskin & Baskin reprennent les travaux de Nikolaeva.
Marianna G. Nikolaeva a conçu un système de classification de la dormance qui reflète le fait que la dormance est déterminée à la fois par les propriétés morphologiques et physiologiques de la graine (Nikolaeva, 1967, 2004). Sur la base de ce système, Baskin & Baskin (1998, 2004) ont proposé un système de classification complet qui comprend cinq classes de dormance des semences : physiologique (PD), morphologique (MD), morphophysiologique (MPD), physique (PY) et combinatoire (PY + PD). Le système, qui est résumé ci-dessous, est hiérarchique, ces cinq classes étant divisées en niveaux et en types.
Finch-Savage WE, Leubner-Metzger G. Seed dormancy and the control of germination. New Phytol. 2006;171(3):501-23. doi: 10.1111/j.1469-8137.2006.01787.x. PMID: 16866955.
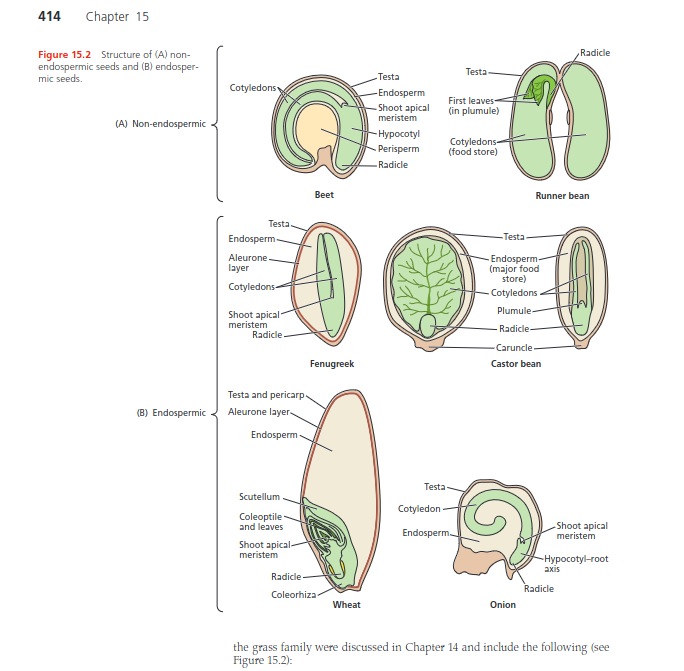
Une graine est l’organe de la plante constituée d’un embryon, de tissus de réserves (albumen ou
endosperme) enfermés dans des enveloppes protectrices (téguments) de morphologies différentes selon
l’espèce (Fenner 2005)1) . L’embryon est constitué de trois parties : la radicule, l’hypocotyle et
l’épicotyle.
Le principal rôle des graines est de fournir une protection et des nutriments à l’embryon durant la
germination (Schmidt 2000)2)
Généralités et considérations
Pour Baskin & Baskin, une graine dormante n’a pas les capacités de germer tant que les facteurs abiotiques sont en dessous du seuil favorable à la germination. Une deuxième raison pour laquelle les graines ne germent pas est qu'une propriété de la graine (ou de l'unité de dispersion) l'empêche. Ainsi, l'absence de germination est un problème lié à la semence plutôt qu'à l'environnement (Eira et Caldas, 2000). La dormance qui résulte d'une caractéristique de la graine est appelée dormance organique (par opposition à dormance imposée) (Nikolaeva, 1969, 1977), et c'est ce type de dormance qui intéresse le plus les biologistes et les écologistes spécialistes des semences.
Par conséquent, la dormance des graines joue un rôle important dans la régulation du moment de la germination afin que les conditions environnementales soient favorables à la survie de la plantule et finalement à la maturation de la plante (Geneve, 2003 ; Finch-Savage et Leubner-Metzger, 2006). Il n'est donc pas surprenant qu'une “caractéristique intéressante de la dormance des graines soit que les plantes ont évolué vers différents mécanismes pour induire la dormance” (Penfield et King, 2009), et aussi pour en sortir.
les graines ne sont pas seulement dormantes pendant la ou les saisons défavorables à la survie des semis, mais les conditions de cette saison peuvent être nécessaires pour rompre la dormance. Ainsi, le stade de la graine dormante dans le cycle de vie de nombreuses espèces végétales doit être considéré comme une période pendant laquelle il se passe des choses dans la graine, par exemple des changements physiologiques/ biochimiques et morphologiques/anatomiques, la croissance de l'embryon, la mobilisation des réserves alimentaires et certainement l'activation et la désactivation de gènes. Étant donné que la levée de la dormance a lieu pendant la saison défavorable à la croissance des semis (ou immédiatement après que l'environnement devient favorable à la croissance des semis dans le cas de nombreuses semences dont l'enveloppe des graines/fruits est imperméable à l'eau), les semences peuvent potentiellement germer au début de la période favorable. Ainsi, les plantules peuvent croître pendant toute la saison favorable.
Baskin & Baskin, 2014
Comme l'expliquent Vleeshouwers et al. (1995), la levée de la dormance et la germination sont deux processus différents et peuvent avoir des exigences différentes. Læ biologiste des semences doit donc prêter attention à la fois à la levée de la dormance et aux exigences de la germination pour réussir à faire germer les semences de nombreuses espèces (Vleeshouwers et al., 1995 ; Srivastava, 2001 ; Thompson et Ooi, 2010).
Une graine est considérée comme ayant germée lorsque sa radicelle fait au moins 2mm de long (Baskin & Baskin 1998). Dans le cas de graines minuscules la graine est comptée comme germée dès la sortie de la radicule. Auparavant, le processus de germination était caractérisé par la cinétique de germination (Baskin & Baskin 1998). Depuis 2005, le processus de germination est défini et mesuré par paramètres
Chez Taiz et Zeiger
Dans Fundamentals of Plant Physiology (2018)
During seed maturation, the embryo dehydrates and enters a quiescent phase. Seed germination requires rehydration and can be defined as the resumption of growth of the embryo in the mature seed. However, the process of germination encompasses all the events that take place between the start of imbibition
(moistening) of the dry seed (discussed later in the context of seed germination) and the emergence of the embryo, usually starting with the radicle, from the structures that surround it. Successful completion of germination depends on the same environmental conditions as vegetative growth (see Chapter 16): Water
and oxygen must be available, and the temperature must be in the physiological
range (that is, the range that does not inhibit physiological processes). However, a viable (living) seed may not germinate even if the appropriate environmental requirements are satisfied, a phenomenon known as seed dormancy. Seed dormancy is an intrinsic temporal block to the initiation of germination that provides additional time for seed dispersal over greater distances. It also maximizes seedling survival by preventing germination under unfavorable conditions.
Avec en ajout :
- Dormance des graines : État d'arrêt de la croissance de l'embryon qui empêche la germination, même lorsque toutes les conditions environnementales nécessaires à la croissance, telles que l'eau, l'oxygène et la température, sont réunies, telles que l'eau, l'Oxygène et la température.
- Quiescence de la graine : État de croissance suspendue de l'embryon en raison d'un manque d'eau, d'eau ou de température adéquate pour la croissance. La germination des graines quiescentes. La germination des graines quiescentes a lieu immédiatement lorsque les conditions évoquées ci-avant sont réunies.
- Dormance primaire L'incapacité des graines matures nouvellement dispersées à germer dans des conditions de croissance normales.
- Dormance secondaire : Les graines qui ont perdu leur dormance primaire peuvent se transformer en graines à croissance rapide ou redevenir dormantes si elles sont exposées de manière prolongée à des conditions de croissance défavorables.
- Dormance imposée par le tégument (ou exogène) : Dormance des graines qui est imposée par l'enveloppe de la graine et d'autres tissus environnants ; elle peut impliquer une imperméabilité à l'eau ou à l'oxygène, une contrainte mécanique ou la rétention de l'extrémité de la graine dans l'enveloppe.
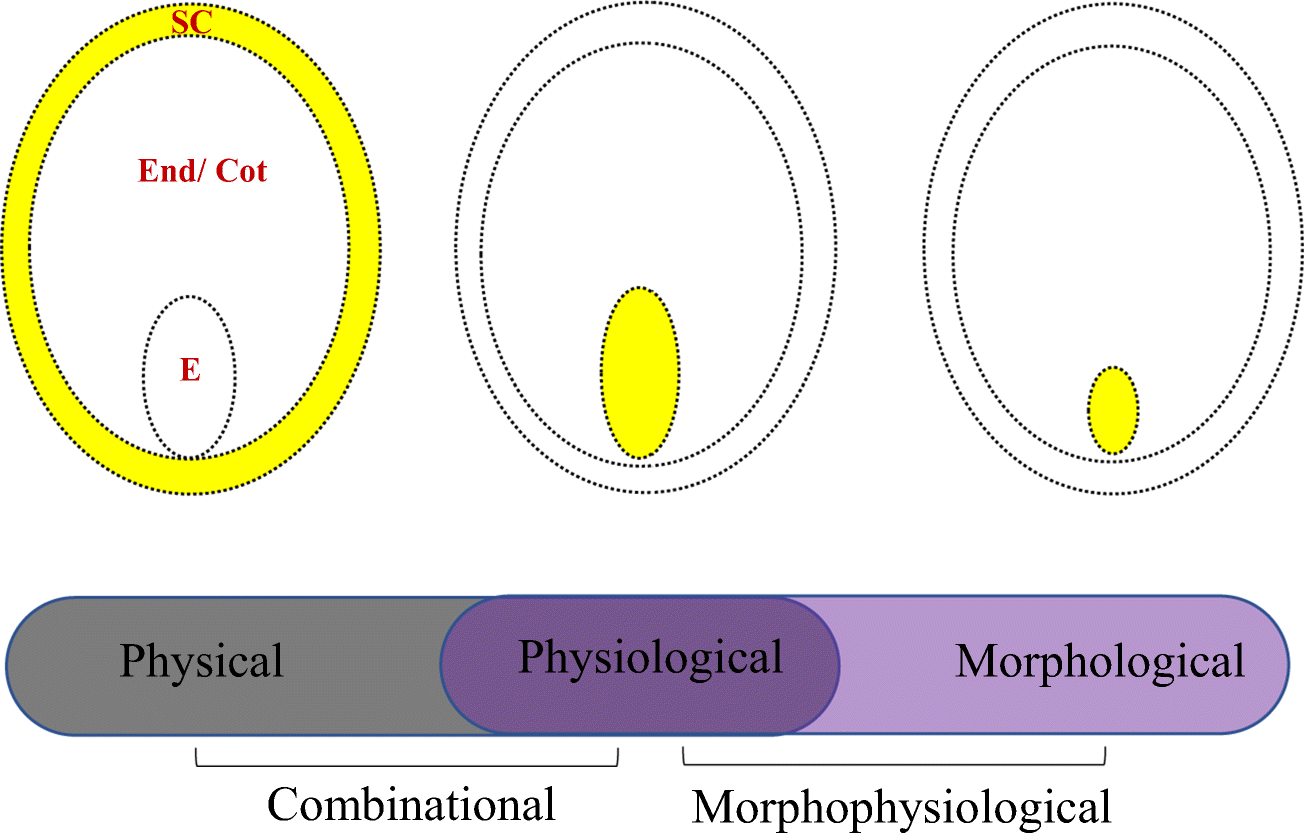
L'angle physiologiste de Taiz et Zeiger peut apparaître plus simpliste que celui de Baskin et Baskin, approches chimiques pour étudier la biologie, qui pourrait sembler plus complet.
Tout cela confirme la complexité, les intrications, la multitude de facteurs et paramètres variants concernant la sortie de dormance puis la germination d'une graine pour une espèces donné, et de toutes la variations entre différentes espèces.
Classification par Baskin et Baskin
 14, 1–16 DOI: 10.1079/SSR2003150")
Dormance primaire
- La dormance physiologique (DPg) est la forme la plus abondante et se retrouve dans les graines de la majorité des angiospermes. Elle met en cause un ou plusieurs mécanismes physiologiques qui proviennent de l’embryon et qui inhibent l’émergence de la radicule (Baskin et Baskin, 1998)5) . Toutefois, les structures qui entourent l’embryon, telles l'albumen ou les téguments, ne sont pas à négliger comme cause potentielle de ce type de dormance. Alors que la dormance morphologique est rattachée à un embryon immature qualifié de rudimentaire ou linéaire, la dormance physiologique n'est liée à aucun critère morphologique et peut se présenter chez n’importe quel type de graines (Nivot 2005)6) .
- La dormance de l'épicotyle est le terme utilisé pour décrire la dormance des graines dans les espèces où il y a un retard d'environ 3-4 semaines (ou plus) dans l'émergence de la pousse après l'émergence de la radicule.
- La dormance morphologique (DM) est due à la présence d’un embryon sous développé en termes de taille (Finch-Savage & Leubner-Metzger 2006). La germination ne peut avoir lieu tant que l’embryon n’est pas arrivé au terme de sa croissance. Avec 2 sous-dormances :
- Dormance physiologique de l'épicotyle : La dormance épicotyle est le terme utilisé pour décrire la dormance des graines dans les espèces où il y a un retard d'environ 3-4 semaines (ou plus) dans l'émergence de la pousse après l'émergence de la radicule.
- La dormance morpho-physiologique (DMPg) qui combine la dormance morphologique et physiologique (Baskin & Baskin 2004). Avec
- dormance morphophysiologique de l'épicotyle (MPD)
- La dormance physique-physiologique (DPqPg) qui associe dormance tégumentaire et dormance physiologique (Nikolaeva 1977 ; Baskin & Baskin 2001, 2004). La germination ne peut se produire que si les deux types de dormance sont levés à la fois. Pour certaines espèces la dormance physique est levée avant la dormance physiologique et réciproquement pour d’autres (Finch-Savage & Leubner-Metzger 2006)9) .
Jaganathan, G.K. Defining correct dormancy class matters: morphological and morphophysiological dormancy in Arecaceae. Annals of Forest Science 77, 100 (2020). https://doi.org/10.1007/s13595-020-01010-7
Autres types non inclus par Baskin et Baskin
Classement par dormance endogène et dormance exogène
Endogène
La dormance endogène est causée par des conditions à l'intérieur de l'embryon lui-même, et elle est aussi souvent divisée en trois sous-groupes : la dormance physiologique, la dormance morphologique et la dormance combinée, chacun de ces groupes pouvant également avoir des sous-groupes.
- Dormance physiologique
- Dessiccation : certaines plantes, dont un certain nombre de graminées et celles des régions arides saisonnières, ont besoin d'une période de dessiccation avant de pouvoir germer ; les graines sont libérées, mais leur teneur en eau doit être réduite avant que la germination ne puisse commencer. Si les graines restent humides après leur dispersion, la germination peut être retardée de plusieurs mois, voire de plusieurs années. De nombreuses plantes herbacées des zones climatiques tempérées ont une dormance physiologique qui disparaît avec le séchage des graines.
- La photodormance ou sensibilité à la lumière affecte la germination de certaines graines. Ces graines photoblastiques ont besoin d'une période d'obscurité ou de lumière pour germer. Chez les espèces dont le tégument est mince, la lumière peut pénétrer dans l'embryon dormant. La présence ou l'absence de lumière peut déclencher le processus de germination, inhibant la germination de certaines graines enfouies trop profondément ou d'autres non enfouies dans le sol.
- La thermodormance est la sensibilité des graines à la chaleur ou au froid. Certaines graines, dont la lampourde et l'amarante, ne germent qu'à des températures élevées (30 °C ou 86 °F). De nombreuses plantes dont les graines germent au début ou au milieu de l'été sont thermodormantes et ne germent que lorsque la température du sol est élevée. D'autres graines ont besoin d'un sol frais pour germer, tandis que d'autres encore, comme le céleri, sont inhibées lorsque la température du sol est trop élevée. Souvent, les exigences en matière de thermodormance disparaissent lorsque la graine vieillit ou se dessèche.
- Dormance morphologique
- Dormance combinée_
Ces graines présentent une dormance à la fois morphologique et physiologique.
La dormance morpho-physiologique ou morphophysiologique se produit lorsque les graines dont les embryons sont sous-développés présentent également des composantes physiologiques de la dormance. Ces graines nécessitent donc des traitements de levée de dormance ainsi qu'une période de temps pour développer des embryons pleinement développés.
- Intermédiaire simple
- Simple profond
- Épicotyle simple profond
- Double simple profond
- Complexe intermédiaire
- Complexe profond
Exogène
La dormance exogène est causée par des conditions extérieures à l'embryon et est souvent divisée en trois sous-groupes :
- La dormance physique
- est le résultat d'une ou de plusieurs couches imperméables qui se développent pendant la maturation et le séchage de la graine ou du fruit
- Dormance mécanique
- se produit lorsque les téguments des graines ou d'autres couvertures sont trop durs pour permettre à l'embryon de se développer pendant la germination[25]. Dans le passé, ce mécanisme de dormance était attribué à un certain nombre d'espèces qui se sont avérées avoir des facteurs endogènes pour leur dormance à la place. Ces facteurs endogènes comprennent un faible potentiel de croissance de l'embryon.
- Dormance chimique
- Comprend les régulateurs de croissance, etc., qui sont présents dans les enveloppes autour de l'embryon. Ils peuvent être éliminés des tissus par lavage ou trempage de la graine, ou désactivés par d'autres moyens. D'autres produits chimiques qui empêchent la germination sont éliminés des graines par l'eau de pluie ou la fonte des neiges.

Le classement de Nikolaeva (1977) comprend :
- 3 types
- Dormance exogène [A]
- Physique
- Chimique
- Mécanique
- Dormance endogène
- Physiologique [C]
- Morphologique [B]
- Morpho-physiologique [B-C]
- Mixte exogène et endogène [A-B-C]
Baskin & Baskin classe ainsi
- 2 divisions
- Division 1. Dormance Imposé [forcé, quiescent] (nondormant, semence ne germe pas en raison de l'absence de conditions environnementales favorables, y compris la lumière) ; pas de sous-catégories
- Division 2. Dormance Organique [innée] (dormante, la graine ne germe pas en raison des propriétés des graines elles-mêmes)
- Subdivision 1. Exogène [A]
- Classe 1. Physique [Aph] (pas de sous-catégories, mais voir Baskin & Baskin, 2004)
- Subdivision 2. Endogène [B et C]
- Classe 2. Morphologique [B] (une seule catégorie)
- Classe 3. Physiologique [C]
- Subclass 1. Regular
- Level 1. Nondeep (C1)a
- Type 1. C 1b or C 1a
- Type 2. C 1b or C 1a
- Type 3. C 1b or C 1a
- Type 4. C 1b
- Type 5. C 1b or C 1a
- Level 2. Intermediate (C2)
- Type 1. (C2b) [not confirmed]
- Type 2. (C2a )
- Level 3. Deep (C3 )
- Type 1. (C3b)
- Type 2. (C3a )
- Subclass 2. Epicotyl
- Level 1. Nondeep
- Type 1. C nd(root)-Cp01b (shoot)
- Type 2. C nd(root)-Cp1b (shoot)
- Type 3. C u1b (root)-Cu1b (shoot)
- Level 2. Deepb,c,d
- Type 1. C nd(root)-C3a(shoot)
- Type 2. C 1b(root)-Cp03b (shoot)
- Type 3. C nd(root)-Cp3b (shoot)
- Type 4.Cu1b (root)-Cu3b (shoot)
- Class 4. Morphophysiological [B-C]
- Subclass 1. Simple
- Level 1. Nondeep (C1Bb)
- Type 1. (C1bBb) (C1bB-C1b sensu Nikolaeva)
- Type 2. (C1a Bb) (C1a B-C1b sensu Nikolaeva)
- Level 2. Intermediate (C1bBb-C2a ) e
- Level 3. Deep (C1b Bb-C3a )e
- Level 4. Nondeep epicotylb,f
- Type 1. C ndBb(root)-C1b(shoot)
- Type 2. C 1bBb(root)-C1b(shoot)
- Type 3. C 1bBb(root)-C1b B0b(shoot)
- Type 4. C 1bBb(root)-C1a (shoot)
- Level 5. Deep epicotyl f,g
- Type 1. C 3bBb(root)-C3b(shoot)
- Type 2. C 1bBb(root)-C3a (shoot)
- Type 3. C 1bBb(root)-C3 B0a (shoot)
- Type 4. C 1bBb(root)-C3a Ba (shoot)
- Level 6. Deep simple double
- Type 1. C 3aBb” (root)-Ce3a (shoot)
- Type 2. C 3aBb0 (root)-Ch3a (shoot)
- Subclass 2. Complexe
- Level 1. Nondeep (C1b1aBa-C 1a)
- Level 2. Intermediate (C2Ba -C2a )
- Level 3. Deep (C3a Ba -C3a )
- Class 5. Combinational [A-C]
- Level 1. Nondeep (Aph-C1)
- Type 1. (Aph-C1b)
- Type 2. (Aph-C1a )
- Level 2. Intermediate (Aph-C2) [not confirmed]
- Type 1 (Aph -C2b)
- Type 2 (Aph -C2a)
- Level 3. Deep (Aph-C 3) [not confirmed]
- Type 1 (Aph -C3b)
- Type 2 (Aph -C3a)
Cette classification fait l'objet de discussions et d'efforts de précisions par actualisation12) 13) 14)
Les options de sortie et/ou rupture de dormance
Graines, dormance et perspectives dans les bioessais
Questionnements
Philosophique et politique
Stratégique
Méthodologique
Notes et références
Sec. Plant Development and EvoDevo. Volume 8 - 2017 https://doi.org/10.3389/fpls.2017.00524